1.0 Spectrum and Wavelengths of Sunlight
Sunlight is a form of electromagnetic energy. Practically about 43% of the total radiant energy emitted from the sun is in the visible spectrum while the remainder lies in the near-infrared (49%) and ultraviolet section (7%).1 However, only less than 1% of the solar radiation is emitted as X-rays, γ-waves and radio waves (Figure 1). The amount of energy delivered is directly proportional to the frequency of the radiation and, is in turn, inversely related to the wavelengths.
 Figure 1.Electromagnetic spectrum of sunlight. Source: http://www.ucar.edu/learn/1_3_1.htm
Figure 1.Electromagnetic spectrum of sunlight. Source: http://www.ucar.edu/learn/1_3_1.htm
1.1 Effective Penetration of Sunlight
As these rays penetrate the Earth’s atmosphere, some of them will be scattered, absorbed, refracted or reflected back into space. The stratosphere has a layer of ozone that absorbs UV radiation2 while the troposphere contains clouds and suspended particles that help to reflect and scatter some of these rays. The portion of light (usually the visible spectrum) that can be captured and used by phototrophs for photosynthesis is called the Photosynthetically Available Radiation.3
As sunlight penetrates the water surfaces, almost 50% of the radiation will be absorbed. Absorption is greatest for both the longer (red, orange) and shorter (UV, violet) wavelengths of light but somewhat less so for the middle range of wavelengths (blue, green) (Figure 2).4 The depth in the water column to which light penetrates is also determined by the extent to which it is absorbed and scattered by dissolved compounds and suspended particles contained within the water. As a general rule, light intensity declines exponentially with depth as described from the Beer-Lambert equation.5 Thus beyond certain depth, insufficient light penetration will impede photosynthesis.
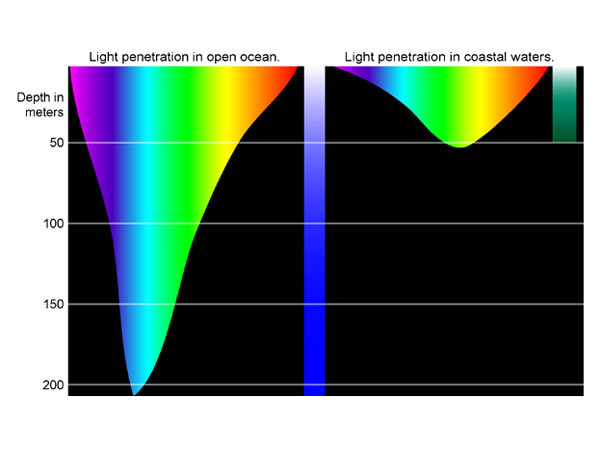 Figure 2: Light penetration at different depths of ocean waters. Source: http://oceanexplorer.noaa.gov/explorations/04deepscope/background/
Figure 2: Light penetration at different depths of ocean waters. Source: http://oceanexplorer.noaa.gov/explorations/04deepscope/background/deeplight/media/diagram3.html
1.0 Photosynthetic Pigments
It is through photosynthesis that full use of solar energy can be harnessed by phototrophs for reducing CO2 to carbohydrates for biomass production. However the productivity of such requires the presence of photosynthetic pigments. In general pigments are molecules that absorb specific wavelengths of light and impart colour from the wavelengths of light reflected.
Biological pigments are usually non-covalently attached to proteins (except for chlorosomes)6 to form pigment-protein complexes which are organized as the photosynthetic unit (PSU). Bacterial PSU consists of 2 types of pigment-protein complexes: the photosynthetic reaction centre (RC) and the light harvesting complexes (LHCs).7 The RC is the exact locale where photosynthetic reaction occurs while the LHCs function merely as antenna that harvest light energy to be channelled to RC for photosynthesis (Figure 3).8
The protein molecules impose an appropriate molecular geometry on the photosynthetic pigments, binding them close together with respect to one another for efficient transfer of energy.9
 Figure 3. The accessory (antenna) pigments transfer the light energy absorbed to adjacent pigments in the LHC until the energy is trapped in the RC. Source: http://fig.cox.miami.edu/~cmallery/150/phts/phts.htm
Figure 3. The accessory (antenna) pigments transfer the light energy absorbed to adjacent pigments in the LHC until the energy is trapped in the RC. Source: http://fig.cox.miami.edu/~cmallery/150/phts/phts.htm2.1 Chlorophylls and Bacteriochlorophlls (Bchls)
Chlorophyll is a greenish pigment that contains a porphyrin ring (Mg at the centre) that is attached to a long hydrophobic phytol tail. The pigment is architecturally designed in such a way that the phytol tail acts to embed the chlorophylls to the lipid and hydrophobic proteins of the membrane while exposing the porphyrin ring outside of it for the absorption of light energy.10 The π-electron system present on the porphyrin ring permits the delocalisation of electrons and could thus provide energised electrons to other molecules. All photosynthetic organisms contain either chlorophyll a (algae and cyanobacteria) or Bchl a (photosynthetic bacteria) as the main pigment in RC. However, depending on the types of organisms, various forms of chlorophylls and Bchls exist that act merely as accessory pigments in the LHCs.
2.2 Carotenoids
Carotenoids are hydrophobic pigments with a conjugated double-bond system and exist in lipid membranes. They are usually yellow or red and absorb light in the blue region of the spectrum. Carotenoids mainly function as accessory pigments that capture light energy not absorbed by the chlorophylls and transfer it to the RC. Besides that carotenoids may also have photoprotective role and is usually associated with chlorophyll. This is due to the fact that excess intense sunlight often always induce the formation of triplet state in chlorophyll, that, when react with oxygen, will form singlet oxygen that could cause cellular damages. Thus carotenoids function to quench this singlet oxygen.11 Since phototrophs must by their very nature live in the light, the photoprotective role of carotenoids is thus an obvious advantage.
2.3 Phycobiliproteins and Phycobilisomes
The phycobiliproteins (phycoerythrin, phycocyanin and allophycocyanin) are composed of a number of subunits, each having a protein backbone to which prosthetic chromophores (phycocyanobilin, phycoerythrobilin, phycourobilin, cryptoviolin) are bound via thioether linkages (Figure 4). Phycobiliproteins are the major light-harvesting pigments in cyanobacteria and red algae and are present as aggregates called phycobilisomes in the cytoplasm or the stroma of chloroplasts.12
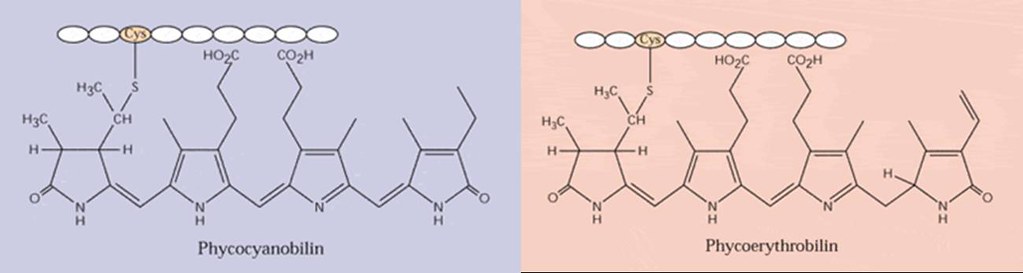
Figure 4. Molecular structures of phycobiliproteins.
The phycobilisomes are constructed in such a way that the pigments are arranged according to their spectral forms, with those that fluoresce at the shortest wavelengths (highest energy) at the tips of the rods (phycoerythrin, 550 nm) while those at the longest wavelengths (lowest energy), at the core end of the rods (phycocyanin, 620 nm). Energy transfer within the rods is thus from the tips to the core, that is, down the energy gradient. Eventually the transferred energy will be channelled to allophycocyanin (650 nm) and ultimately to Bchl a on the thylakoid membrane (Figure 5).13


Figure 5. Rod-like structures of phycobilisomes. Sources: http://hypnea.botany.uwc.ac.za/phylogeny/classif/cyan2.htm, http://www.external.ameslab.gov/news/Inquiry/fall95/light.html
1.0 Range of absorptions
Absorption spectrum is a plot of the amount of light absorbed across a series of wavelengths while the action spectrum is a measure of the rate of photosynthesis as a function of wavelengths of light. The specific preference of absorption for certain wavelengths was initially shown by Engelmann in that filamentous green alga illuminated with a tiny spectrum of visible light was surrounded by aerotactic bacteria around the portions of red and blue light – showing the maximum absorptions of chlorophyll (Figure 6).14 For photosynthesis, the action spectrum is very similar to the absorption spectra of chlorophylls (Figure 7).
The fact that each pigment only absorbs strongly at certain specific wavelengths (Figures 8), the efficient use of the broad spectrum of sunlight emitted will be a waste if there is no mean of harvesting all of them together. Therefore to compensate for that, (bacterio)chlorohylls exist not just only in one form but in several variants. Moreover phototrophs contain several other accessory pigments (carotenoids, xanthophylls, phycobilins) in addition to (bacterio)chlorophylls to harvest light energy at wavelengths over which the latter absorbs poorly.

Figure 6. Engelman’s experiment. Source: http://www.plantphys.net/article.php?ch=7&id=66
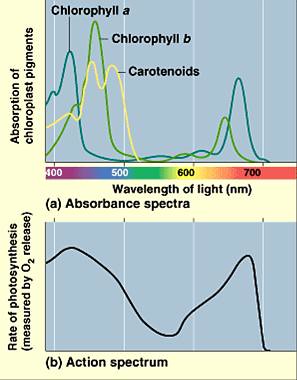
Figure 7. Absorption spectra and action spectrum of photosynthesis. Source:
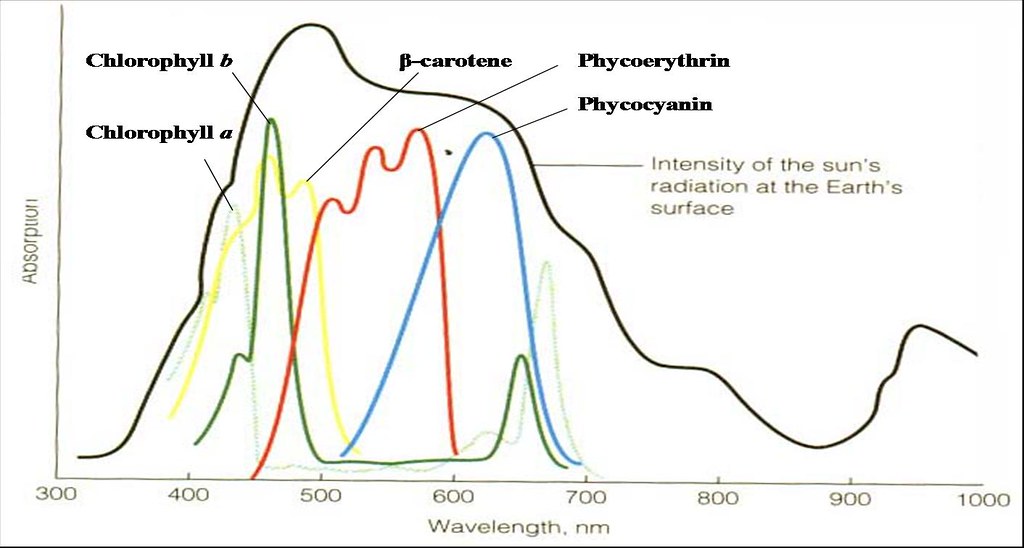
Figure 8. Absorption spectra of chlorophylls and other accessory pigments.
Pigment | Chl a | Chl b | Bchl a | Bchl b | β-carotene | Phyco-erythrin | Phyco-cyanin |
| Absorption maximum (nm) | 430, 680 | 463, 660 | 364, 770 | 373, 795 | 450 | 550 | 620 |
4.0 Photosynthetic (micro)organisms
Photosynthetic (micro)organisms consist mainly of algae and bacteria and are further divided based on their photosynthetic pathways – oxygenic photosynthesis (algae, cyanobacteria), anaerobic anoxygenic photosynthesis (AnAnP) and aerobic anoxygenic photosynthesis (AAnP).15 In oxygenic photosynthesis, electrons are removed from H2O to form O2. Anoxygenic photosynthesis on the other hand, use light energy to extract electrons from molecules other then water, and thus O2 is not evolved.16 The difference between AnAnp and AAnP is that the latter requires O2 while the former demands on anaerobiosis, but both equally do not generate O2 as the by-product of photosynthesis.
In contrast to the presence of chloroplasts in the eukaryotic algae, prokaryotes do not contain one, and the photosynthetic pigments are either integrated into the internal membrane systems that arise from the invagination of the plasma membrane (purple bacteria), the plasma membrane itself (heliobacteria), in both the plasma membrane and specialized non-unit membrane-enclosed structures called chlorosomes (green bacteria) or in the thylakoid membranes (cyanobacteria).10
| Algae | Bacteria |
| Chlorophyta (green algae) Charophyta (stoneworts / brittleworts) Euglenophyta (euglenoids) Chrysophyta (goled-brown, yellow-green algae, diatoms) Phaeophyta (brown algae) Rhodophyta (red algae) Pyrrhophyta (dinoflagellates) | Cyanobacteria and prochlorophytes (phylum Cyanobacteria) Purple sulphur bacteria (γ-proteobacteria - families Chromatiaceae and Ectothiorhodospiraceae) Purple non-sulphur bacteria (α-proteobacteria – 5 families; β-proteobacteria – 1 family) Green sulphur bacteria (phylum Chlorobi) Green non-sulphur bacteria (phylum Chloroflexi) Heliobacteria. |
5.0 Factors influencing Vertical Distribution of Phototrophs
In the aquatic environment, particularly the oceans where most phototrophs are aplenty, the rate of photosynthesis is limited by the intensity and spectral composition of light that penetrates the water surface. In general photosynthesis increases with light intensity until it becomes light saturated, beyond which, photoinhibition will ensue (Figure 11).17 Nevertheless light is quickly attenuated and selectively absorbed as it passes through water. Thus the ability of phototrophs to utilise available light energy depends largely upon their ability to absorb the available wavelengths, which, in turn, is determined by their photosynthetic pigment content (Figure 9), that overall, influence the vertical distribution of phototrophic microorganisms (Figure 10).18,19
For instance, the green and euglenoid algae contain chlorophylls a and b; diatoms, dinoflagellates and brown algae contain chlorophylls a, c and special carotenoids while red algae and cyanobacteria contain chlorophyll a and phycobilins.5 The content of phycobilisome is able to increase as light intensity decreases and thus allows growth at fairly low light intensities, a feature termed chromatic adaptation.20 Since blue light penetrates the deepest into water column, the energy of wavelengths greater than 600 nm to which chlrorophyll absorbs maximally is thus a selective advantage.

Figure 9. Absorption spectra of algae and cyanobacteria with complementary use of light by sulphur bacteria. Source: http://jan.ucc.nau.edu/~doetqp-p/
courses/env440/env440_2/lectures/lec23/lec23.html

Figure 10. Widnogradsky column. Source: http://jan.ucc.nau.edu/~doetqp-p/courses/env440/env440_2/lectures/lec23/lec23.html
Phytoplanktons such as cyanobacteria and algae are usually confined to the euphotic zone where the availability of sunlight is the greatest. The requirement for dissolved O2 and atmospheric N2 among certain cyanobacteria, also impose an overall need to remain suspended on water surfaces. Nevertheless the density (0.999-1.26 g cm-3) with which phytoplankton is generally associated often suggested a natural tendency for them to sink.21 However the elongated features of most algae and the extensive projections of diatoms provide an increased surface area that confers maximum drag that decreases the sinking velocity. Some phytoplankton however, contains gas vacuoles or flagella to remain afloat. Apart from that, they may also be carried to the top periodically by the turbulence mixing of water column due to wind or storms. 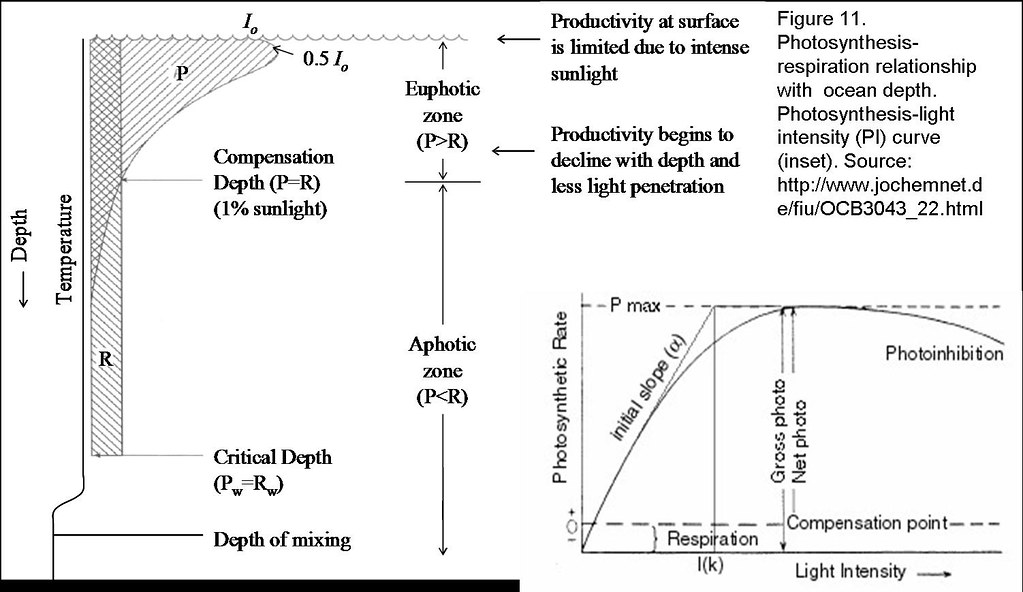
The mixing of phytoplankton throughout the euphotic zone causes them to see a light environment that changes continually. While photosynthesis is light-dependent, respiration is however unaffected by light and remains constant with depth. The depth at which the amount of photosynthesis equals respiration is known as the compensation depth. It represents the minimum amount of photosynthesis just to maintain cellular metabolism. However, phytoplanktons rarely stay at the same depth in the water column. If they are above the compensation depth, they receive full light availability and produce more organic material than is respired. In the deeper epilimnion where they are mixed below the compensation level but above the critical depth, net positive growth will still be observed. However no net growth is noted at the critical depth, beyond with, the phytoplanktons will respire more carbon than they fix, and will soon die. Here, the critical depth refers to the level in which phytoplankton can be mixed and still meet their metabolic requirements from photosynthesis (Figure 11).22, 23
The purple non-sulfur and purple sulfur bacteria on the other hand, usually inhabit water column beneath the phytoplanktons. They make use of the infrared rays (>800 nm) and light of wavelength 500 nm that are transmitted by the sand matrix and not absorbed by phytoplanktons for photosynthesis. This is mediated by the long wavelength absorption of Bchl-protein complexes (B800 and B850 Bchls, absorbing at 800 and 850 nm, respectively) and carotenoids that absorb at 500 nm.24 The photosynthetic apparatus contains a RC that is located in the middle of the ring shaped LHC-I, which is collectively in turn, surrounded by multiple copies of LHC-II (Figure 15). In the LHC-II, both the B800 and B850 (ring structure) Bchls are oriented perpendicular to each other to maximize light absorption from every direction (Figure 14).25, 26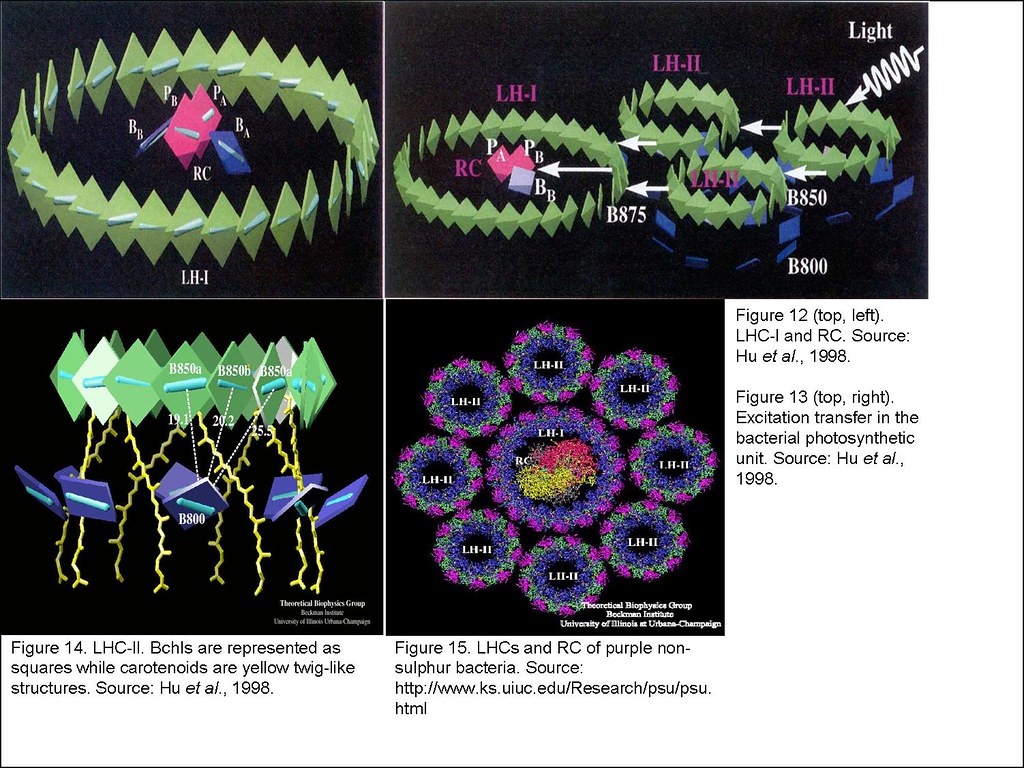
Depending on the availability of light, some purple bacteria are able to grow by respiration in the dark. For instance, the purple non-sulphur bacteria are extremely flexible about their choice of energy source. They normally grow anaerobically as photoorganoheterotrophs. However, in the absence of light they can grow aerobically as chemoorganoheterotrophs. In fact, oxygen inhibits Bchl and carotenoid synthesis, so cultures growing aerobically in the dark are colourless. Because of their metabolism, they are present in the mud and water of lakes and ponds with abundant organic matter. Since the purple non-sulphur bacteria only tolerate low S2- concentrations, they are usually vertically distributed above the purple sulphur bacteria that thrive mainly on the H2S generated by SO42- reducing bacteria in the anoxic sediment.
The green sulfur bacteria are obligate anoxygenic photolithoautotrophs that utilise H2S, S and H2 as electron donors. Collectively, they are found together with the purple sulphur bacteria in the profundal zone whereby their oxidized products, SO42- will be reduced by Desulfovibrio at the benthic zone into H2S to be used latter by both the bacteria when it diffuses upwards. However the green sulphur bacteria are mainly distributed beneath the purple sulphur bacteria due to their high affinity for S2- and their preference for anaerobiosis.27
In addition to that, green sulphur bacteria contain mainly Bchls c, d or e and carotenoids such as chlorobactene and isorenieratene, with which the purple bacteria are not associated with. The LHCs of these bacteria are carried in ellipsoidal vesicles called chlorosomes. The Bchls in the chlorosomes are not associated with proteins but instead function much like a solid state circuit that is efficient in absorbing extreme low light intensity. Moreover, as the light intensity decreases, the size of the Bchl c antenna increases to compensate for light deficit.28 Thus green sulfur bacteria can grow at the lowest light intensities of any known phototrophs. Although they lack flagella and are nonmotile, some species have gas vesicles to adjust their depth for optimal light and H2S.29 Those forms without vesicles are found in S2- rich muds at the bottom of lakes and ponds.
References
- http://www.ucar.edu/learn/1_3_1.htm
- http://www.weatheroffice.pyr.ec.gc.ca/skywatchers/teachersguide/tg_chap06_e.html
- http://www.niwa.cri.nz/pubs/wa/09-1/sunlight.htm
- http://oceanexplorer.noaa.gov/explorations/04deepscope/background/deeplight/media/diagram3.html
- Fletcher, M. 1979. Microbial ecology: a conceptual approach. In The aquatic environment, ed. Lynch, J.M. and
- http://www.personal.psu.edu/faculty/n/x/nxf10/phd/
- http://www.ks.uiuc.edu/Research/psu/psu.html
- http://photoscience.la.asu.edu/photosyn/education/photointro.html
- http://nist.rcsb.org/pdb/molecules/pdb22_1.html
- Madigan, M.T., Martinko, J.M. and Parker, J. 2000. Brock Biology of Microorganims.
- http://www.chem.ufl.edu/~reu/main/projects/Angerhofer.html
- http://www.sbsp.jp/sbsp/Sb/sb41/017.html
- http://www.external.ameslab.gov/news/Inquiry/fall95/light.html
- http://www.plantphys.net/article.php?ch=7&id=66
- Karl, D.M. 2002. Hidden in a sea of microbes. Nature 415: 590-591.
- http://www.purlife.com/photosynthesis.htm
- http://www.jochemnet.de/fiu/OCB3043_22.html
- http://jan.ucc.nau.edu/~doetqp-p/courses/env440/env440_2/lectures/lec23/lec23.html
- http://www.personal.psu.edu/faculty/j/e/jel5/biofilms/winogradsky.html
- Grossman, A.R., Schaefer, M.R., Chiang, G.G. and Collier, J.L. 1993. The phycobilisome, a light-harvesting complex responsive to environmental conditions. Microbiological Reviews 57(3): 725-749.
- http://www.esf.edu/efb/schulz/Limnology/phytoecol1.html
- http://www.jochemnet.de/fiu/OCB3043_22.html
- http://maritime.haifa.ac.il/departm/lessons/ocean/lect26.htm
- Hu, X. and Schulten, K. 1997 (August). How nature harvests sunlight. Physics Today: 28-34.
- Hu, X., Ritz, T., Damjanović, A. and Schulten, K. 1997. Pigment organization and transfer of electronic excitation in the photosynthetic unit of purple bacteria. J. Phys. Chem. B 101: 3854-3871.
- Hu, X., Damjanović, A., Ritz, T. and Schulten, K. 1998. Architecture and mechanism of the light-harvesting apparatus of purple bacteria. Proceedings of the
- http://141.150.157.117:8080/prokPUB/chaphtm/323/11_02.htm
- Yakovlev, A.G., Taisova, A.S. and Fetisova, Z.G. 2002. Light control over the size of an antenna unit building block as an efficient strategy for light harvesting in photosynthesis. FEBS Letters 512: 129-132.
- http://141.150.157.117:8080/prokPUB/chaphtm/323/11_00.htm